Also see:
Exercise and Effect on Thyroid Hormone
Ray Peat, PhD: Quotes Relating to Exercise
Temperature and Pulse Basics & Monthly Log
Ray Peat, PhD on Thyroid, Temperature, Pulse, and TSH
Body Temperature, Metabolism, and Obesity
Thyroid, Temperature, Pulse
Metabolism, Brain Size, and Lifespan in Mammals
Promoters of Efficient v. Inefficient Metabolism
Inflammation from Decrease in Body Temperature
Melatonin Lowers Body Temperature
Menopausal Estrogen Therapy Lowers Body Temperature
Thyroid Function, Pulse Rate, & Temperature
“Curing” a High Metabolic Rate with Unsaturated Fats
Fat Deficient Animals – Activity of Cytochrome Oxidase
Comparison: Carbon Dioxide v. Lactic Acid
Carbon Dioxide Basics
Energy Flow: Plant World and Animal World
Biological Energy & Matter Cycle
Is 98.6 Really Normal?
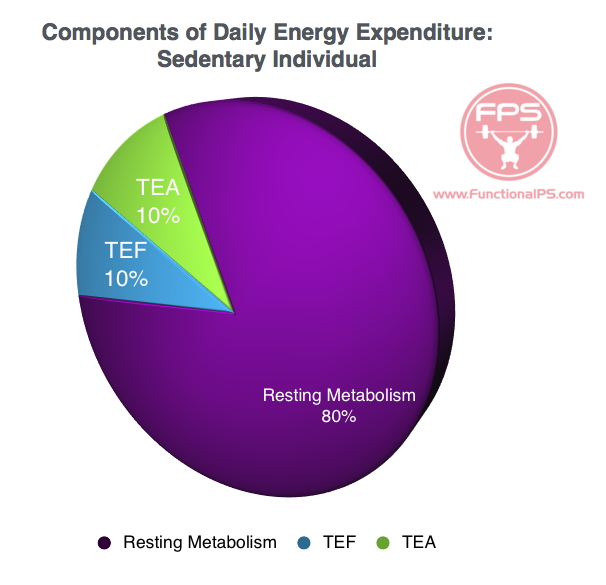
Components of Daily Energy Expenditure
According to Poehlman (1989), daily energy expenditure is comprised of three major components:
1) resting metabolic rate (RMR) — thyroid hormone, lean body mass ~56-80%
2) the thermic effect of feeding (TEF) — food choice ~10+%
3) the thermic effect of activity (TEA) — movement ~10+%
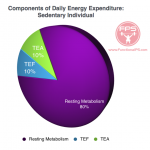
Daily Energy Expenditure – Sedentary
RMR accounts for the largest portion of daily energy expenditure no matter whether you’re sedentary or very active — 80 to 56% respectively (Landsberg, et al., 2009). Energy expenditure from exercise/activity is thus small relative to the energy expenditure at rest. On a percentage basis, more bang for your buck comes from analyzing and maximizing the factors that improve the RMR, with special attention to thyroid hormone and the preservation of lean body mass.
How strong is your resting metabolism? A good way to get a picture of your resting metabolism is by charting your resting temperature and pulse on a daily basis. Temperatures below 98F degrees and pulses not between 75-85 beats per minute are a sign of depressed energy metabolism. Using a finger oximeter can provide instant feedback about resting pulse (and Sp02), making charting more convenient.
Exercise, when used correctly (resistance-training centered, training not draining), can enhance or maintain the RMR through preservation of fat free mass without thyroid suppression. Resistance training does not significantly raise non-resting energy expenditure, but its benefits on weight management come about by “its effects on body composition (e.g., increasing fat-free mass)” (Poehlman & Melby, 1998). The preservation of lean mass with aging helps prevent the accumulation of body fat by sustaining the resting metabolic rate and acting as a site that consumes stored fat at rest.
Three quotes by Ray Peat, PhD on muscle, exercise, and fat loss:
“In the resting state, muscles consume mainly fats, so maintaining relatively large muscles is important for preventing the accumulation of fats.”
“Since fat has a very low rate of metabolism, people who lose muscle by fasting are going to have increasing difficulty in losing weight, since they will have less active tissue to consume fat. Building up muscle and lymph tissue for optimal health – even if it initially causes a slight weight gain – will make reducing easier by increasing mass of metabolically active tissue.”
“Many dietitians claim that exercise doesn’t increase the need for protein, but the Russians have found that a combination of exercise and increased protein intake can increase the muscle mass. In a woman, this process can not only improve grace and body proportions, but it also increases the body’s ability to burn up fat. Other nutrients are needed for using protein properly, and for maintaining optimum nerve functioning. However, if the exercise produces too much stress and not enough muscle action, muscle will atrophy as a result of cortisone’s shifting amino acid metabolism into glucose production.”
When exercise is used incorrectly, the loss of fat free mass and suppression thyroid hormone production results in lowering of the RMR, creating poor long-term outcomes from a weight management perspective. Calorie restrictive diets combined with exercise also suppress the RMR, increasing the likelihood of regaining any lost weight.
Protein is a macronutrient identified in the literature as having a beneficial effect on lean mass preservation (Poehlman, et al., 1993). Since ingestion of adequate carbohydrate prevents protein from being use to make glucose, it too has such an effect. According to Ray Peat, PhD, proteolytic enzymes that digest protein are inhibit by polyunsaturated fats (PUFA). PUFA are also antagonistic the thyroid in multiple ways. Eliminate PUFA from the diet in favor of saturated fats.
Quotables about RMR:
1. “Hence, the age-related decline in BMR is partly explained by a reduction in the quantity, as well as the metabolic activity, of DEXA-derived lean tissue components.” -Piers, et al., 1998
2. “The hypothesis is that combining diet and exercise will accelerate fat loss, preserve fat-free weight and prevent or decelerate the decline in resting metabolic rate more effectively than with diet restriction alone. The optimal combination of diet and exercise, however, remains elusive. It appears that the combination of a large quantity of aerobic exercise with a very low calorie diet resulting in substantial loss of bodyweight may actually accelerate the decline in resting metabolic rate. These findings may cause us to re-examine the quantity of exercise and diet needed to achieve optimal fat loss and preservation of resting metabolic rate.” -Poehlman, et al., 1991
3. “Evidence is presented to suggest that although resistance training may elevate resting metabolic rate, it does not substantially enhance daily energy expenditure in free-living individuals…Preliminary evidence suggests that although resistance training increases muscular strength and endurance, its effects on energy balance and regulation of body weight appear to be primarily mediated by its effects on body composition (e.g., increasing fat-free mass) rather than by the direct energy costs of the resistance exercise.” -Poehlman & Melby, 1998
4. “Results showed that body composition did not change in endurance-trained women, but maximum aerobic capacity increased by 18%. Resistance-trained women increased muscular strength and fat-free mass (1.3 kg).” -Poehlman, et al., 2002
5. “Daily energy expenditure is composed of three major components: 1) resting metabolic rate (RMR); 2) the thermic effect of feeding (TEF); and 3) the thermic effect of activity (TEA). RMR constitutes 60 to 75% of daily energy expenditureand is the energy associated with the maintenance of major body functions. TEF is the cumulative increase in energy expenditure after several meals and constitutes approximately 10% of daily energy expenditure.” -Poehlman, 1989
6. “The direct effects of physical activity interventions on energy expenditure are relatively small when placed in the context of total daily energy demands. Hence, the suggestion has been made that exercise produces energetic benefits in other components of the daily energy budget, thus generating a net effect on energy balance much greater than the direct energy cost of the exercise alone. Resting metabolic rate (RMR) is the largest component of the daily energy budget in most human societies and, therefore, any increases in RMR in response to exercise interventions are potentially of great importance…Long-term effects of training include increases in RMR due to increases in lean muscle mass. Extreme interventions, however, may induce reductions in RMR, in spite of the increased lean tissue mass, similar to the changes observed in animals in response to flight.” -Speakman & Selman, 2003
7. “This review reveals that there might be contrasting effects of resting and nonresting energy expenditure on aging, with increases in the former being protective and increases in the latter being harmful.” -Speakman, et al., 2002
8. “We considered the association of several metabolic and lifestyle variables as modulators of the decline in resting metabolic rate (RMR) and fat-free weight (FFW) in 183 healthy females (18-81 yr). RMR showed a curvilinear decline with age, which was significant in women aged 51-81 yr but not in women aged 18-50 yr. FFW Wshowed a curvilinear decline with age, which was significant (P < 0.01) in women 48-81 yr but not in women 18-47 yr. The decline in RMR was primarily associated with the loss of FFW (r2 = 72%), whereas the decline in FFW was explained primarily by differences in maximal O2 consumption (VO2max), age, leisure time physical activity, and dietary protein intake (total r2 = 46%). We conclude that RMR and FFW showed a curvilinear decline with age which was accelerated beyond the middle-age years. Second, the age-related decline in RMR was primarily associated with the loss of FFW. Third, the loss of FFW was partially related to a decrement in VO2max and nutritional factors.” -Poehlman, et al., 1993
9. “An important goal during weight loss is to maximize fat loss while preserving metabolically active fat-free mass (FFM). Massive weight loss typically results in substantial loss of FFM potentially slowing metabolic rate…Despite relative preservation of FFM, exercise did not prevent dramatic slowing of resting metabolism out of proportion to weight loss. This metabolic adaptation may persist during weight maintenance and predispose to weight regain unless high levels of physical activity or caloric restriction are maintained.” -Johannsen, et al., 2012
10. ” The extremely high failure rate (> 80%) to keep the reduced weight after successful weight loss is due to adaptation processes of the body to maintain body energy stores. This so called “adaptive thermogenesis” is defined as a smaller than predicted change of energy expenditure in response to changes in energy balance. Adaptive thermogenesis appears to be a major reason for weight regain. The foremost objective of weight-loss programs is the reduction in body fat. However, a concomitant decline in lean tissue can frequently be observed. Since lean body mass (LBM) represents a key determinant of RMR it follows that a decrease in lean tissue could counteract the progress of weight loss. Therefore, with respect to long-term effectiveness of weight reduction programs, the loss of fat mass while maintaining LBM and RMR seems desirable” -Weck, et al., 2012
11. “The foremost objective of a weight-loss trial has to be the reduction in body fat leading to a decrease in risk factors for metabolic syndrome. However, a concomitant decline in lean tissue can frequently be observed. Given that fat-free mass (FFM) represents a key determinant of the magnitude of resting metabolic rate (RMR), it follows that a decrease in lean tissue could hinder the progress of weight loss. Therefore, with respect to long-term effectiveness of weight-loss programmes, the loss of fat mass while maintaining FFM and RMR seems desirable…The advantages of strength training may have greater implications than initially proposed with respect to decreasing percentage body fat and sustaining FFM. Research to date suggests that the addition of exercise programmes to dietary restriction can promote more favourable changes in body composition than diet or physical activity on its own…These outcomes provide the scientific rationale to justify further randomised intervention trials on the synergies between diet and exercise approaches to yield favourable modifications in body composition.” -Stiegler & Cunliffe, 2006
11. “Declines in energy expenditure favoring the regain of lost weight persist well beyond the period of dynamic weight loss.” -Rosenbaum, et al., 2008
12. “After 2 weeks of energy restriction, measured REE had fallen by 469 and 635 kJ/d more than predicted and this difference reached 963 and 614 kJ/d by week 8 of treatment in men and women respectively.” -Doucet, et al., 2001
13. “Changes in body temperature are associated with significant changes in metabolic rate.
…
“Resting (or “basal”) metabolic rate (RMR) accounts for approximately 80% of energy output. About two thirds of RMR is for maintenance of homeothermy (warm-bloodedness); about one third is to maintain cellular integrity, ionic gradients, protein turnover, and the like [6-8]. Resting metabolic rate is largely regulated by thyroid hormones, with a minor contribution from the sympathetic nervous system. Resting metabolic rate differs by as much as 600 kcal/d for a 70-kg man [8]. Physical activity (exercise) accounts for about 10% in truly sedentary humans; in addition to intentional activity, this category includes nonpurposeful motion such as fidgeting, which may differ among lean and obese individuals [9], as well as upright posture [10]. The remaining 10% is frequently referred to as thermogenesis, which means heat production unrelated to physical activity.”
It should be emphasized that, for nonsedentary individuals, the activity component may be much greater than 10% of total energy expenditure. Evidence has been developed indicating that the combination of activity plus adaptive thermogenesis accounts for about 44% of total energy expenditure on average, meaning that RMR would constitute about 56% of total energy expenditure in normally active humans [15], as compared with 80% in the truly sedentary.” -Landsberg, et al., 2009
=============================================================
Am J Clin Nutr. 1992 Apr;55(4):802-10.
The effects of either high-intensity resistance or endurance training on resting metabolic rate.
Broeder CE, Burrhus KA, Svanevik LS, Wilmore JH.
The effects of either 12-wk of high-intensity endurance or resistance training on resting metabolic rate (RMR) were investigated in 47 males aged 18-35 y. Subjects were randomly assigned to either a control (C), resistance-trained (RT) or endurance-trained (ET) group. After training both exercise groups showed significant declines in relative body fat either by reducing their total fat weight and maintaining fat-free weight (ET) or by reducing their total fat weight and increasing fat-free weight (RT). RMR did not significantly change after either training regimen although a small decline in energy intake was observed along with an increase in energy expenditure [ET, 2.721 MJ (650 kcal) per training day]. These results suggest that both endurance and resistance training may help to prevent an attenuation in RMR normally observed during extended periods of negative energy balance (energy intake less than expenditure) by either preserving or increasing a person’s fat-free weight.
Journal of Applied Physiology December 1, 1998 vol. 85 no. 6 2196-2204
Is there evidence for an age-related reduction in metabolic rate?
Leonard S. Piers1, Mario J. Soares2, Leanne M. McCormack3, and Kerin O’Dea1
To determine whether the age-related reduction in basal metabolic rate (BMR) is explained by a quantitative and/or qualitative change in the components of lean tissue, we conducted a cross-sectional study in groups of young (n = 38, 18–35 yr) and older (n = 24, 50–77 yr) healthy individuals. BMR was measured by indirect calorimetry. Body composition was obtained by using dual-energy X-ray absorptiometry (DEXA), which permitted four compartments to be quantified [bone mineral mass, fat mass (FM), appendicular lean tissue mass (ALTM), and nonappendicular lean tissue mass (NALTM)]. Absolute BMR and ALTM were lower, whereas FM was significantly higher in the older, compared with young, subjects. BMR, adjusted for differences in FM, ALTM, and NALTM, was significantly lower in the older subjects by 644 kJ/day. In separate regression analyses of BMR on body compartments, older subjects had significantly lower regression coefficients for ALTMand NALTM, compared with young subjects. Hence, the age-related decline in BMR is partly explained by a reduction in the quantity, as well as the metabolic activity, of DEXA-derived lean tissue components.
Sports Med. 1991 Feb;11(2):78-101.
The impact of exercise and diet restriction on daily energy expenditure.
Poehlman ET, Melby CL, Goran MI.
In addition to the direct energy cost of physical activity, exercise may influence resting energy expenditure in 3 ways: (a) a prolonged increase in postexercise metabolic rate from an acute exercise challenge; (b) a chronic increase in resting metabolic rate associated with exercise training; and (c) a possible increase in energy expenditure during nonexercising time. It seems apparent that the greater the exercise perturbation, the greater the magnitude of the increase in postexercise metabolic rate. An exercise prescription for the general population that consists of exercise of low (less than 50% VO2max) or moderate intensity (50 to 75% VO2max) does not appear to produce a prolonged elevation of postexercise metabolic rate that would influence body-weight. Inconsistent results have been found with respect to the effects of exercise training and the trained state on resting metabolic rate. Whereas some investigators have found a higher resting metabolic rate in trained than untrained individuals and in individuals after an exercise training programme, other investigators have found no chronic exercise effect on resting metabolic rate. Differences in experimental design, genetic variation and alterations in energy balance may contribute to the discrepant findings among investigators. A relatively unexplored area concerns the influence of exercise training on energy expenditure during nonexercising time. It is presently unclear whether exercise training increases or decreases the energy expenditure associated with spontaneous or nonpurposeful physical activity which includes fidgeting, muscular activity, etc. The doubly labelled water technique represents a methodological advance in this area and permits the determination of total daily energy expenditure. Concomitant with the determination of the other components of daily energy expenditure (resting metabolic rate and thermic effect of a meal), it will now be possible to examine the adaptive changes in energy expenditure during nonexercising time. A plethora of studies have examined the combined effects of diet and exercise on body composition and resting metabolic rate. The hypothesis is that combining diet and exercise will accelerate fat loss, preserve fat-free weight and prevent or decelerate the decline in resting metabolic rate more effectively than with diet restriction alone. The optimal combination of diet and exercise, however, remains elusive. It appears that the combination of a large quantity of aerobic exercise with a very low calorie diet resulting in substantial loss of bodyweight may actually accelerate the decline in resting metabolic rate. These findings may cause us to re-examine the quantity of exercise and diet needed to achieve optimal fat loss and preservation of resting metabolic rate.
Int J Sport Nutr. 1998 Jun;8(2):143-59.
Resistance training and energy balance.
Poehlman ET, Melby C.
In this brief review we examine the effects of resistance training on energy expenditure. The components of daily energy expenditure are described, and methods of measuring daily energy expenditure are discussed. Cross-sectional and exercise intervention studies are examined with respect to their effects on resting metabolic rate, physical activity energy expenditure, postexercise oxygen consumption, and substrate oxidation in younger and older individuals. Evidence is presented to suggest that although resistance training may elevate resting metabolic rate, it does not substantially enhance daily energy expenditure in free-living individuals. Several studies indicate that intense resistance exercise increases postexercise oxygen consumption and shifts substrate oxidation toward a greater reliance on fat oxidation. Preliminary evidence suggests that although resistance training increases muscular strength and endurance, its effects on energy balance and regulation of body weight appear to be primarily mediated by its effects on body composition (e.g., increasing fat-free mass) rather than by the direct energy costs of the resistance exercise.
J Clin Endocrinol Metab. 2002 Mar;87(3):1004-9.
Effects of endurance and resistance training on total daily energy expenditure in young women: a controlled randomized trial.
Poehlman ET, Denino WF, Beckett T, Kinaman KA, Dionne IJ, Dvorak R, Ades PA.
There exists considerable controversy regarding the impact of different modes of exercise training on total daily energy expenditure (TEE). To examine this question, young, nonobese women were randomly assigned to a supervised 6-month program of endurance training, resistance training, or control condition. TEE was measured before and 10 d after a 6-month exercise program was completed with doubly labeled water. Body composition was determined from dual energy x-ray absorptiometry, maximum aerobic capacity from a treadmill test to exhaustion, and muscular strength from one-repetition maximum tests. Results showed that body composition did not change in endurance-trained women, but maximum aerobic capacity increased by 18%. Resistance-trained women increased muscular strength and fat-free mass (1.3 kg). TEE did not significantly change when measured subsequent to the endurance or resistance training programs. Absolute resting metabolic rate increased in resistance-trained women but not when adjusted for fat-free mass. No change in physical activity energy expenditure was found in any of the groups. These results suggest that endurance and resistance training does not chronically alter TEE in free-living young women. Thus, the energy-enhancing benefits of exercise training are primarily derived from the direct energy cost of exercise and not from a chronic elevation in daily energy expenditure in young, nonobese women.
Med Sci Sports Exerc. 1989 Oct;21(5):515-25.
A review: exercise and its influence on resting energy metabolism in man.
Poehlman ET.
Daily energy expenditure is composed of three major components: 1) resting metabolic rate (RMR); 2) the thermic effect of feeding (TEF); and 3) the thermic effect of activity (TEA). RMR constitutes 60 to 75% of daily energy expenditure and is the energy associated with the maintenance of major body functions. TEF is the cumulative increase in energy expenditure after several meals and constitutes approximately 10% of daily energy expenditure. Most investigators, however, have examined the thermic effect of a single meal test (TEM). TEA is the most variable component of daily energy expenditure and can constitute 15 to 30% of 24-h energy expenditure. This component includes energy expenditure due to physical work, muscular activity, including shivering and fidgeting, as well as purposeful physical exercise. Participation in purposeful exercise (both acute and chronic) is a subcomponent of TEA and has been found to influence resting energy expenditure (RMR and TEM). Reports in the literature, however, are discrepant regarding the direction and magnitude of the effects of exercise and exercise training on RMR and TEM. Cross-sectional and longitudinal studies that have examined the effects of exercise on RMR and TEM are reviewed. Possible explanations for divergent results in the literature are discussed. The major focus of this review is directed to human studies, although pertinent animal work is included. The role of genetic variation, gender specific responses, and methodological considerations for future studies examining the relation among RMR, TEA, and TEM are considered. Although still controversial, purposeful physical exercise appears to influence resting energy expenditure in man.
Proc Nutr Soc. 2003 Aug;62(3):621-34.
Physical activity and resting metabolic rate.
Speakman JR, Selman C.
The direct effects of physical activity interventions on energy expenditure are relatively small when placed in the context of total daily energy demands. Hence, the suggestion has been made that exercise produces energetic benefits in other components of the daily energy budget, thus generating a net effect on energy balance much greater than the direct energy cost of the exercise alone. Resting metabolic rate (RMR) is the largest component of the daily energy budget in most human societies and, therefore, any increases in RMR in response to exercise interventions are potentially of great importance. Animal studies have generally shown that single exercise events and longer-term training produce increases in RMR. This effect is observed in longer-term interventions despite parallel decreases in body mass and fat mass. Flight is an exception, as both single flights and long-term flight training induce reductions in RMR. Studies in animals that measure the effect of voluntary exercise regimens on RMR are less commonly performed and do not show the same response as that to forced exercise. In particular, they indicate that exercise does not induce elevations in RMR. Many studies of human subjects indicate a short-term elevation in RMR in response to single exercise events (generally termed the excess post-exercise O2 consumption; EPOC). This EPOC appears to have two phases, one lasting < 2 h and a smaller much more prolonged effect lasting up to 48 h. Many studies have shown that long-term training increases RMR, but many other studies have failed to find such effects. Data concerning long-term effects of training are potentially confounded by some studies not leaving sufficient time after the last exercise bout for the termination of the long-term EPOC. Long-term effects of training include increases in RMR due to increases in lean muscle mass. Extreme interventions, however, may induce reductions in RMR, in spite of the increased lean tissue mass, similar to the changes observed in animals in response to flight.
J Nutr. 2002 Jun;132(6 Suppl 2):1583S-97S.
Living fast, dying when? The link between aging and energetics.
Speakman JR, Selman C, McLaren JS, Harper EJ.
The idea that aging should be linked to energy expenditure has a long history that can be traced to the late 1800s and the industrial revolution. Machines that are run fast wear out more quickly, so the notion was born that humans and animals might experience similar fates: the faster they live (expressed as greater energy expenditure), the sooner they die. Evidence supporting the “rate-of-living” theory was gleaned from the scaling of resting metabolism and life span as functions of body mass. The product of these factors yields a mass-invariant term, equivalent to the “amount of living.” There are at least four problems with this evidence, which are summarized and reviewed in this communication: 1) life span is a poor measure of aging, 2) resting metabolism is a poor measure of energy expenditure, 3) the effects are confounded by body mass and 4) the comparisons made are not phylogenetically independent. We demonstrate that there is a poor association between resting metabolic rate (RMR) and daily energy expenditure (DEE) measured using the doubly labeled water (DLW) method at the level of species. Nevertheless, the scaling relation between DEE and body mass still has the same scaling exponent as the RMR and body mass relationship. Thus, if we use DEE rather than RMR in the analysis, the rate-of-living ideas are still supported. Data for 13 species of small mammal were obtained, where energy demands by DLW and longevity were reliably known. In these species, there was a strong negative relationship between residual longevity and residual DEE, both with the effects of body mass removed (r(2) = 0.763, F = 32.1, P < 0.001). Hence, the association of energy demands and life span is not attributed to the confounding effects of body size. We subjected these latter data to an analysis that extracts phylogenetically independent contrasts, and the relationship remained significant (r(2) = 0.815, F = 39.74, P < 0.001). Small mammals that live fast really do die young. However, there are very large differences between species in the amounts of living that each enjoy and these disparities are even greater when other taxa are included in the comparisons. Such differences are incompatible with the “rate-of-living” theory. However, the link between energetics and aging across species is reconcilable within the framework of the “free-radical damage hypothesis” and the “disposable soma hypothesis.” Within species one might anticipate the rate-of-living model would be more appropriate. We reviewed data generated from three different sources to evaluate whether this were so, studies in which metabolic rate is experimentally increased and impacts on life span followed, studies of caloric restriction and studies where links between natural variation in metabolism and life span are sought. This review reveals that there might be contrasting effects of resting and nonresting energy expenditure on aging, with increases in the former being protective and increases in the latter being harmful.
AJP – Endo March 1, 1993 vol. 264 no. 3 E450-E455
Determinants of decline in resting metabolic rate in aging females
E. T. Poehlman, M. I. Goran, A. W. Gardner, P. A. Ades, P. J. Arciero, S. M. Katzman-Rooks, S. M. Montgomery, M. J. Toth, and P. T. Sutherland
We considered the association of several metabolic and lifestyle variables as modulators of the decline in resting metabolic rate (RMR) and fat-free weight (FFW) in 183 healthy females (18-81 yr). RMR showed a curvilinear decline with age, which was significant in women aged 51-81 yr but not in women aged 18-50 yr. FFW Wshowed a curvilinear decline with age, which was significant (P < 0.01) in women 48-81 yr but not in women 18-47 yr. The decline in RMR was primarily associated with the loss of FFW (r2 = 72%), whereas the decline in FFW was explained primarily by differences in maximal O2 consumption (VO2max), age, leisure time physical activity, and dietary protein intake (total r2 = 46%). We conclude that RMR and FFW showed a curvilinear decline with age which was accelerated beyond the middle-age years. Second, the age-related decline in RMR was primarily associated with the loss of FFW. Third, the loss of FFW was partially related to a decrement in VO2max and nutritional factors. Therapeutic interventions designed to increase VO2max by elevating physical activity may preserve fat-free weight and thus offset the decline of RMR in aging women.
J Clin Endocrinol Metab. 2012 Jul;97(7):2489-96. doi: 10.1210/jc.2012-1444. Epub 2012 Apr 24.
Metabolic slowing with massive weight loss despite preservation of fat-free mass.
Johannsen DL, Knuth ND, Huizenga R, Rood JC, Ravussin E, Hall KD.
CONTEXT:
An important goal during weight loss is to maximize fat loss while preserving metabolically active fat-free mass (FFM). Massive weight loss typically results in substantial loss of FFM potentially slowing metabolic rate.
OBJECTIVE:
Our objective was to determine whether a weight loss program consisting of diet restriction and vigorous exercise helped to preserve FFM and maintain resting metabolic rate (RMR).
PARTICIPANTS AND INTERVENTION:
We measured body composition by dual-energy x-ray absorptiometry, RMR by indirect calorimetry, and total energy expenditure by doubly labeled water at baseline (n = 16), wk 6 (n = 11), and wk 30 (n = 16).
RESULTS:
At baseline, participants were severely obese (× ± SD; body mass index 49.4 ± 9.4 kg/m(2)) with 49 ± 5% body fat. At wk 30, more than one third of initial body weight was lost (-38 ± 9%) and consisted of 17 ± 8% from FFM and 83 ± 8% from fat. RMR declined out of proportion to the decrease in body mass, demonstrating a substantial metabolic adaptation (-244 ± 231 and -504 ± 171 kcal/d at wk 6 and 30, respectively, P < 0.01). Energy expenditure attributed to physical activity increased by 10.2 ± 5.1 kcal/kg.d at wk 6 and 6.0 ± 4.1 kcal/kg.d at wk 30 (P < 0.001 vs. zero).
CONCLUSIONS:
Despite relative preservation of FFM, exercise did not prevent dramatic slowing of resting metabolism out of proportion to weight loss. This metabolic adaptation may persist during weight maintenance and predispose to weight regain unless high levels of physical activity or caloric restriction are maintained.
Dtsch Med Wochenschr. 2012 Oct;137(43):2223-8. doi: 10.1055/s-0032-1327232. Epub 2012 Oct 17.
[Strategies for successful weight reduction – focus on energy balance].
[Article in German]
Weck M, Bornstein SR, Barthel A, Blüher M.
The prevalence of obesity and related health problems is increasing worldwide and also in Germany. It is well known that substantial and sustained weight loss is difficult to accomplish. Therefore, a variety of studies has been performed in order to specify causes for weight gain and create hypotheses for better treatment options. Key factors of this problem are an adaptation of energy metabolism, especially resting metabolic rate (RMR), non-exercise thermogenesis and diet induced thermogenesis. The extremely high failure rate (> 80%) to keep the reduced weight after successful weight loss is due to adaptation processes of the body to maintain body energy stores. This so called “adaptive thermogenesis” is defined as a smaller than predicted change of energy expenditure in response to changes in energy balance. Adaptive thermogenesis appears to be a major reason for weight regain. The foremost objective of weight-loss programs is the reduction in body fat. However, a concomitant decline in lean tissue can frequently be observed. Since lean body mass (LBM) represents a key determinant of RMR it follows that a decrease in lean tissue could counteract the progress of weight loss. Therefore, with respect to long-term effectiveness of weight reduction programs, the loss of fat mass while maintaining LBM and RMR seems desirable. In this paper we will discuss the mechanisms of adaptive thermogenesis and develop therapeutic strategies with respect to avoiding weight regain successful weight reduction.
Sports Med. 2006;36(3):239-62.
The role of diet and exercise for the maintenance of fat-free mass and resting metabolic rate during weight loss.
Stiegler P, Cunliffe A.
The incidence of obesity is increasing rapidly. Research efforts for effective treatment strategies still focus on diet and exercise programmes, the individual components of which have been investigated in intervention trials in order to determine the most effective recommendations for sustained changes in bodyweight. The foremost objective of a weight-loss trial has to be the reduction in body fat leading to a decrease in risk factors for metabolic syndrome. However, a concomitant decline in lean tissue can frequently be observed. Given that fat-free mass (FFM) represents a key determinant of the magnitude of resting metabolic rate (RMR), it follows that a decrease in lean tissue could hinder the progress of weight loss. Therefore, with respect to long-term effectiveness of weight-loss programmes, the loss of fat mass while maintaining FFM and RMR seems desirable. Diet intervention studies suggest spontaneous losses in bodyweight following low-fat diets, and current data on a reduction of the carbohydrate-to-protein ratio of the diet show promising outcomes. Exercise training is associated with an increase in energy expenditure, thus promoting changes in body composition and bodyweight while keeping dietary intake constant. The advantages of strength training may have greater implications than initially proposed with respect to decreasing percentage body fat and sustaining FFM. Research to date suggests that the addition of exercise programmes to dietary restriction can promote more favourable changes in body composition than diet or physical activity on its own. Moreover, recent research indicates that the macronutrient content of the energy-restricted diet may influence body compositional alterations following exercise regimens. Protein emerges as an important factor for the maintenance of or increase in FFM induced by exercise training. Changes in RMR can only partly be accounted for by alterations in respiring tissues, and other yet-undefined mechanisms have to be explored. These outcomes provide the scientific rationale to justify further randomised intervention trials on the synergies between diet and exercise approaches to yield favourable modifications in body composition.
Br J Nutr. 2009 Aug;102(4):488-92. doi: 10.1017/S0007114508207245.
Adaptive reduction in thermogenesis and resistance to lose fat in obese men.
Tremblay A, Chaput JP.
Adaptive thermogenesis is defined as a greater than predicted change in energy expenditure in response to changes in energy balance. This issue is particularly relevant in the context of a weight-reducing programme in which diminished thermogenesis can be sufficient to compensate for a prescribed decrease in daily energy intake. In the present pilot study, we investigated the adaptive reduction in thermogenesis in resting state that appears to favour resistance to further weight loss. Eight obese men (mean BMI: 33.4 kg/m2, mean age: 38 years) participated in this repeated-measures, within-subject, clinical intervention. They were subjected to a weight-loss programme that consisted of a supervised diet (-2930 kJ/d) and exercise clinical intervention. The phases investigated were as follows: (i) baseline, (ii) after 5 (SE 1) kg loss of body weight (phase 1), (iii) after 10 (SE 1) kg weight loss (phase 2) and (iv) at resistance to further weight loss (plateau). At each phase of the weight-reducing programme, body weight and composition as well as RMR were measured. A regression equation was established in a control population of the same age to predict RMR in obese men at each phase of the weight-loss programme. We observed that body weight and fat mass (FM) were significantly reduced (P < 0.05), while fat-free mass remained unchanged throughout the programme. In phase 1, measured RMR had fallen by 418 kJ/d, more than predicted (P < 0.05), and this difference reached 706 kJ/d at plateau (P < 0.05 v. phase 1). A positive association (r 0.64, P < 0.05) was observed between the reduction in thermogenesis and the degree of FM depletion at plateau. The adaptive reduction in thermogenesis at plateau was substantial and represented 30.9% of the compensation in energy balance that led to resistance to further lose body weight. In conclusion, these results show that adaptive reduction in thermogenesis may contribute to the occurrence of resistance to lose fat in obese men subjected to a weight-reducing programme.
Am J Clin Nutr. 2008 Oct;88(4):906-12.
Long-term persistence of adaptive thermogenesis in subjects who have maintained a reduced body weight.
Rosenbaum M, Hirsch J, Gallagher DA, Leibel RL.
BACKGROUND:
After weight loss, total energy expenditure — in particular, energy expenditure at low levels of physical activity — is lower than predicted by actual changes in body weight and composition. An important clinical issue is whether this reduction, which predisposes to weight regain, persists over time.
OBJECTIVE:
We aimed to determine whether this disproportionate reduction in energy expenditure persists in persons who have maintained a body-weight reduction of > or =10% for >1 y.
DESIGN:
Seven trios of sex- and weight-matched subjects were studied in an in-patient setting while receiving a weight-maintaining liquid formula diet of identical composition. Each trio consisted of a subject at usual weight (Wt(initial)), a subject maintaining a weight reduction of > or =10% after recent (5-8 wk) completion of weight loss (Wt(loss-recent)), and a subject who had maintained a documented reduction in body weight of >10% for >1 y (Wt(loss-sustained)). Twenty-four-hour total energy expenditure (TEE) was assessed by precise titration of fed calories of a liquid formula diet necessary to maintain body weight. Resting energy expenditure (REE) and the thermic effect of feeding (TEF) were measured by indirect calorimetry. Nonresting energy expenditure (NREE) was calculated as NREE = TEE – (REE +TEF).
RESULTS:
TEE, NREE, and (to a lesser extent) REE were significantly lower in the Wt(loss-sustained) and Wt(loss-recent) groups than in the Wt(initial) group. Differences from the Wt(initial) group in energy expenditure were qualitatively and quantitatively similar after recent and sustained weight loss.
CONCLUSION:
Declines in energy expenditure favoring the regain of lost weight persist well beyond the period of dynamic weight loss.
Br J Nutr. 2001 Jun;85(6):715-23.
Evidence for the existence of adaptive thermogenesis during weight loss.
Doucet E, St-Pierre S, Alméras N, Després JP, Bouchard C, Tremblay A.
The present study was performed to further investigate the adaptive component of thermogenesis that appears during prolonged energy restriction. Fifteen obese men and twenty obese women underwent a 15-week weight-loss programme. During this programme, body weight and composition as well as resting energy expenditure (REE) were measured at baseline, after 2 and 8 weeks of energy restriction (-2929 kJ/d) and drug therapy (or placebo), and finally 2-4 weeks after the end of the 15-week drug therapy and energy restriction intervention, when subjects were weight stable. Regression equations were established in a control population of the same age. These equations were then used to predict REE in obese men and women at baseline, after 2 and 8 weeks, as well as after the completion of the programme. In both men and women body weight and fat mass were significantly reduced in all cases) while fat-free mass remained unchanged throughout the programme. At baseline, REE predicted from the regression equation was not significantly different from the measured REE in men, while in women the measured REE was 13 % greater than predicted. After 2 weeks of energy restriction, measured REE had fallen by 469 and 635 kJ/d more than predicted and this difference reached 963 and 614 kJ/d by week 8 of treatment in men and women respectively. Once body-weight stability was recovered at the end of the programme, changes in REE remained below predicted changes in men (-622 kJ/d). However, in women changes in predicted and measured REE were no longer different at this time, even if the women were maintaining a reduced body weight. In summary, the present results confirm the existence of adaptive thermogenesis and give objective measurements of this component during weight loss in obese men and women, while they also emphasize that in women this component seems to be essentially explained by the energy restriction.
Metabolism. 2009 Jun;58(6):871-6. doi: 10.1016/j.metabol.2009.02.017.
Is obesity associated with lower body temperatures? Core temperature: a forgotten variable in energy balance. (full paper)
Landsberg L, Young JB, Leonard WR, Linsenmeier RA, Turek FW.
The global increase in obesity, along with the associated adverse health consequences, has heightened interest in the fundamental causes of excessive weight gain. Attributing obesity to “gluttony and sloth”, blaming the obese for overeating and limiting physical activity, oversimplifies a complex problem, since substantial differences in metabolic efficiency between lean and obese have been decisively demonstrated. The underlying physiological basis for these differences have remained poorly understood. The energetic requirements of homeothermy, the maintenance of a constant core temperature in the face of widely divergent external temperatures, accounts for a major portion of daily energy expenditure. Changes in body temperature are associated with significant changes in metabolic rate. These facts raise the interesting possibility that differences in core temperature may play a role in the pathophysiology of obesity. This review explores the hypothesis that lower body temperatures contribute to the enhanced metabolic efficiency of the obese state.
Some quotables:
“Resting (or “basal”) metabolic rate (RMR) accounts for
approximately 80%of energy output. About two thirds of RMR is for maintenance of homeothermy (warm-bloodedness); about one third is to maintain cellular integrity, ionic gradients, protein turnover, and the like [6-8] Resting metabolic rate is largely
regulated by thyroid hormones, with a minor contribution from the sympathetic nervous system. Resting metabolic rate differs by as much as 600 kcal/d for a 70-kg man [8]. Physical activity (exercise) accounts for about 10% in truly sedentary humans; in addition to intentional activity, this category includes nonpurposeful motion such as fidgeting, which may differ among lean and obese individuals [9], as well as upright posture [10]. The remaining 10% is frequently referred to as thermogenesis, which means heat production unrelated to physical activity.”
“It should be emphasized that, for nonsedentary individuals, the activity component may be much greater than 10% of total energy expenditure. Evidence has been developed indicating that the combination of activity plus adaptive thermogenesis accounts for about 44% of total energy expenditure on average, meaning that RMR would constitute about 56% of total energy expenditure in normally active humans [15], as compared with 80% in the truly sedentary.“
 latest posts")